


اطلاعات عمومی، ویکی ژن
آسیب DNA و ترمیم DNA: انواع و مکانیسم
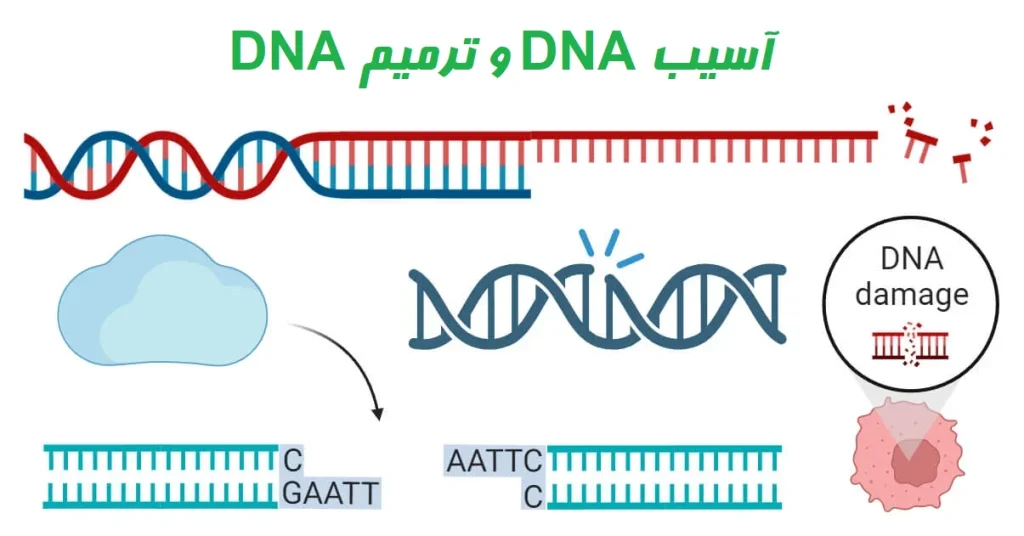
مقدمهای بر آسیب DNA و ترمیم DNA
DNA واحد اصلی وراثت است که یکپارچگی و عملکرد ارگانیسمها را حفظ میکند. با این حال به طور مداوم در معرض عوامل آسیب رسان است که میتواند باعث آسیب دیدن DNA شود. علاوه بر این، خطاهایی در طول فرآیندهای همانندسازی (Replication) و ترمیم DNA رخ میدهد که منجر به ایجاد جهشهای مضر میشود.
برای غلبه بر اثرات مضر آسیب DNA، سلولها دارای سیستمهای مختلفی مانند مکانیسمهای ترمیم DNA، مسیرهای تحمل آسیب، نقاط کنترلی چرخه سلولی و مسیرهای مرگ سلولی هستند. این سیستمها با هم کار میکنند تا آسیب DNA را ترمیم یا تحمل کرده و بقای کلی و کارکرد سلولها را تضمین کنند.
درک مکانیسمهای آسیب DNA و مسیرهای ترمیم مربوطه برای فهم تأثیر آسیب DNA بر عملکرد سلولی و ظهور بیماریها ضروری است.
آسیب DNA چیست؟
آسیب DNA به تغییرات یا اختلالاتی اطلاق میشود که در مولکول DNA اتفاق میافتد و ناشی از عوامل محیطی یا فرآیندهای طبیعی است که در داخل سلولهای ما رخ میدهد.
انواع آسیب DNA و مکانیسمها
انواع مختلفی از آسیب DNA وجود دارند که میتوانند به دلیل فرآیندهای سلولی طبیعی یا قرار گرفتن در معرض عوامل مخرب در محیط اتفاق بیفتند.
تعدادی از انواع آسیب DNA به شرح زیر است:
-
شکستگیهای رشته DNA
شکستگی رشته DNA زمانی اتفاق میافتد که یک یا هر دو رشته DNA قطع شود. دو نوع شکستگی وجود دارد: شکستگیهای تک رشتهای (Single-strand breaks (SSBs)) که در آن یک رشته بریده میشود و شکستگیهای دو رشتهای (Double-strand breaks (DSBs)) که در آن هر دو رشته بریده میشوند. این شکستگیها میتوانند در اثر پرتو یونیزان (Ionizing radiation) مانند اشعه ایکس و گاما و همچنین برخی مواد شیمیایی ایجاد شوند.
-
آسیب اکسیداتیو
آسیب اکسیداتیو میتواند به دلیل فعالیت گونههای فعال اکسیژن (Reactive oxygen species (ROS)) رخ دهد که منجر به تشکیل ضایعاتی میشود. ROS بسیار واکنشپذیر، مانند رادیکالهای هیدروکسیل (OH•) ، میتواند باعث آسیب اکسیداتیو به بازهای DNA شود.
-
آلکیلاسیون بازها (Alkylation of Bases)
عوامل آلکیله کننده، هم درونزا و هم برونزا، میتوانند بازهای DNA را با معرفی گروههای آلکیل تغییر دهند. این تغییرات میتوانند سیتوتوکسیک (Cytotoxic) یا جهشزا (Mutagenic) بوده یا دارای اثرات خنثی بر روی سلول باشند.
-
از دست دادن باز (Base Loss)
از دست دادن باز زمانی رخ میدهد که بازهای نیتروژنی در DNA برداشته شده و سایتهای آپورینیک (Apurinic) / آپریمیدینیک (Apyrimidinic) (که به اختصارسایتهای AP مینامند) یا سایتهای آبازیک (Abasic) باقی بمانند. سایتهای AP از نظر شیمیایی ناپایدار هستند و در صورت عدم ترمیم میتوانند منجر به شکستن رشته DNA یا رویدادهای جهشزا شوند.
-
تشکیل ترکیبات اضافی حجیم (Bulky Adduct)
ترکیبات افزایشی حجیم زمانی تشکیل میشوند که مواد شیمیایی خاصی مانند هیدروکربنهای آروماتیک چند حلقهای (Polycyclic aromatic hydrocarbons (PAHs)) به طور کووالانسی به بازهای DNA متصل شوند. این ترکیبات افزایشی تغییرات حجمی زیادی را ایجاد میکنند که از DNA بیرون زده و ساختار آن را مختل میکنند. آنها میتوانند در فرآیندهای همانندسازی، رونویسی (Transcription) و ترمیم DNA تداخل داشته باشند و به طور بالقوه منجر به جهش شوند.
-
اتصال عرضی DNA
اتصال عرضی DNA زمانی اتفاق میافتد که دو نوکلئوتید (Nucleotide) در DNA به صورت کووالانسی به یکدیگر متصل میشوند. اتصالهای عرضی میتوانند در همان رشته DNA (اتصالهای عرضی درون رشتهای (Intrastrand)) یا بین رشتههای DNA مقابل هم (اتصالهای عرضی بین رشتهای (Interstrand)) ایجاد شوند. اتصالهای عرضی DNA از جدا شدن رشتههای DNA در حین فرایند تکثیر یا رونویسی جلوگیری میکنند و منجر به ایجاد اختلال در فرآیندهای مهم سلولی میشوند.
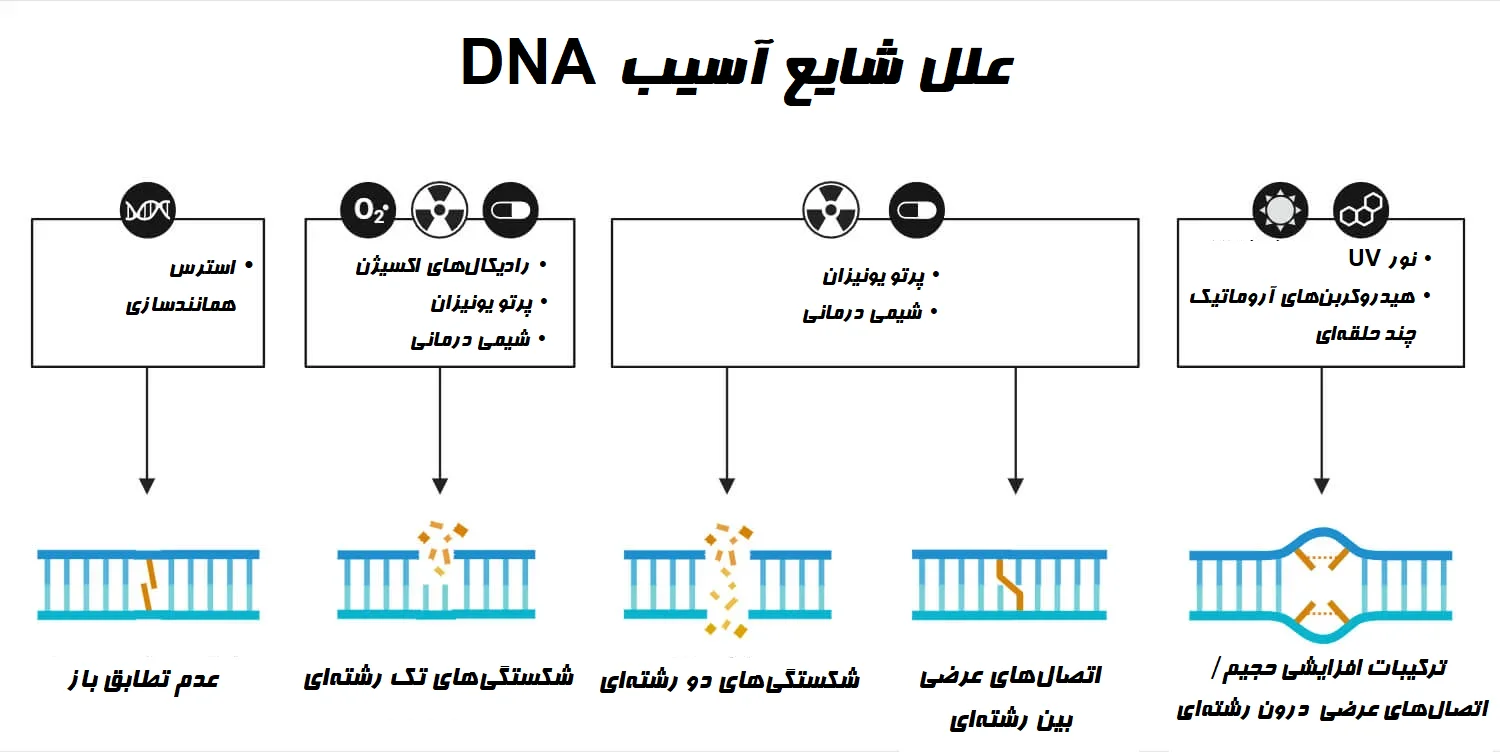
منابع / عوامل آسیب DNA
آسیب DNA را همچنین میتوان بر اساس منشاء یا منابع آن به دو نوع درونزا (Endogenous) و برونزا (Exogenous) طبقهبندی کرد. منابع اصلی آسیب DNA درونزا و برونزا به طور خلاصه در زیر توضیح داده شده است:
-
آسیب DNA درونزا
آسیب DNA درونزا از واکنشهای داخلی شامل DNA از نظر شیمیایی فعال در درون سلولها سرچشمه میگیرد.
- خطاهای همانندسازی یکی از منشاءهای آسیب DNA درونزا هستند که در حین فرایند همانندسازی DNA زمانی که نوکلئوتیدهای نادرست در مقابل بازهای الگو قرار میگیرند، رخ میدهد. در طول همانندسازی، برخی از DNA پلیمرازها با دقت عمل کمتر میتوانند درگیر شوند که منجر به بروز خطاهای بالقوه میشود.
- آنزیمهای توپوایزومراز (Topoisomerase) منبع دیگری از آسیب DNA درونزا هستند. توپوایزومرازها در حین فرایندهای همانندسازی و رونویسی، ابرپیچش DNA را حذف میکنند. با این حال، ناهمترازی انتهای DNA میتواند کمپلکس برش توپوایزومراز-DNA را تثبیت کند و منجر به تشکیل ضایعات DNA شود.
- گونههای فعال اکسیژن (ROS) در طی فرآیندهای سلولی تولید میشوند و میتوانند باعث آسیب اکسیداتیو به DNA شوند. در حالی که ROS نقش مهمی در عملکردهای سلولی طبیعی ایفا میکند، سطوح بیش از حد آن میتواند منجر به ایجاد ضایعات و تغییرات مختلف در DNA شود. ROS بیش از حد با ایجاد چندین بیماری انسانی مانند سرطان، بیماری آلزایمر و دیابت مرتبط است.
- عوامل آلکیله کننده، ترکیبات واکنشی هستند که میتوانند گروههای متیل (Methyl) یا اتیل (Ethyl) را به بازهای DNA اضافه کنند و منجر به ایجاد تغییرات شیمیایی شوند. رویدادهای متیلاسیون (methylation) خود به خودی میتوانند بازهای متیله متفاوتی ایجاد کنند. برخی از بازهای متیله جهشزا هستند و میتوانند منجر به بروز انواع خاصی از جهش شوند.
-
آسیب DNA برونزا
آسیب DNA برونزا توسط عوامل خارجی مانند عوامل محیطی، نیروهای فیزیکی یا مواد شیمیایی ایجاد میشود.
- پرتو یونیزان (IR) مستقیماً به DNA آسیب میزند یا به طور غیرمستقیم از طریق تولید رادیکالهای هیدروکسیل (OH) بسیار واکنش پذیر از مولکولهای آب بر آن تأثیر میگذارد. IR میتواند انواع مختلفی از آسیب به DNA مانند ضایعات بازی و شکستگیهای تک رشتهای و دو رشتهای ایجاد کند.
- اشعه ماوراء بنفش (UV) یکی دیگر از عوامل آسیب DNA است. این اشعه عامل اصلی سرطان پوست است. نور فرابنفش میتواند دایمرهای پیریمیدین (pyrimidine dimer) را ایجاد کند که در آن دو پیریمیدین در یک رشته DNA به هم متصل میشوند. این تغییر در ساختار DNA میتواند مانع فرآیندهای رونویسی و همانندسازی شود.
- عوامل آلکیله کننده برونزا که در منابعی مانند دود تنباکو و فعالیتهای صنعتی یافت میشوند، با DNA واکنش میدهند و میتوانند تغییرات جهشزا و سرطانزا ایجاد کنند. آنها اساساً بازهای نیتروژنی در DNA را هدف قرار میدهند. نمونههایی از عوامل آلکیله کننده شامل سولفور و نیتروژن موستارد (Nitrogen mustard) میباشد.
- آمینهای آروماتیک (Aromatic amine) که در دود سیگار، سوخت، زغالسنگ، رنگها و آفتکشها یافت میشوند نیز منابع برونزای آسیب DNA هستند. این عوامل میتوانند ضایعات طولانی مدتی را در ساختار DNA ایجاد کنند که منجر به جایگزینی بازهای DNA و ایجاد جهشهای دگرقالب (Frameshift mutation) میشوند.
- هیدروکربن های آروماتیک چند حلقهای (Polycyclic aromatic hydrocarbons (PAHs)) مواد سرطانزای شناخته شدهای هستند که در منابعی مانند دود تنباکو، اگزوز خودروها و سایر آلایندههای محیطی یافت میشوند. PAHها نیاز به فعالسازی توسط سیستم P-450 کبدی دارند تا مواد واکنشی تولید کنند که به طور بالقوه میتواند به DNA آسیب برساند.
ترمیم DNA چیست؟
- آسیب DNA یک رویداد رایج است که میتواند با فرآیندهای مهم سلولی تداخل داشته باشد و منجر به بروز نقص ژنومی و افزایش خطر سرطان شود. سلولها برای اطمینان از یکپارچگی ژنوم خود، جهت ایجاد مکانیسمهایی برای ترمیم DNA تکامل یافتهاند. این مکانیسمها به سلولها کمک میکنند تا با آسیب DNA مقابله نمایند.
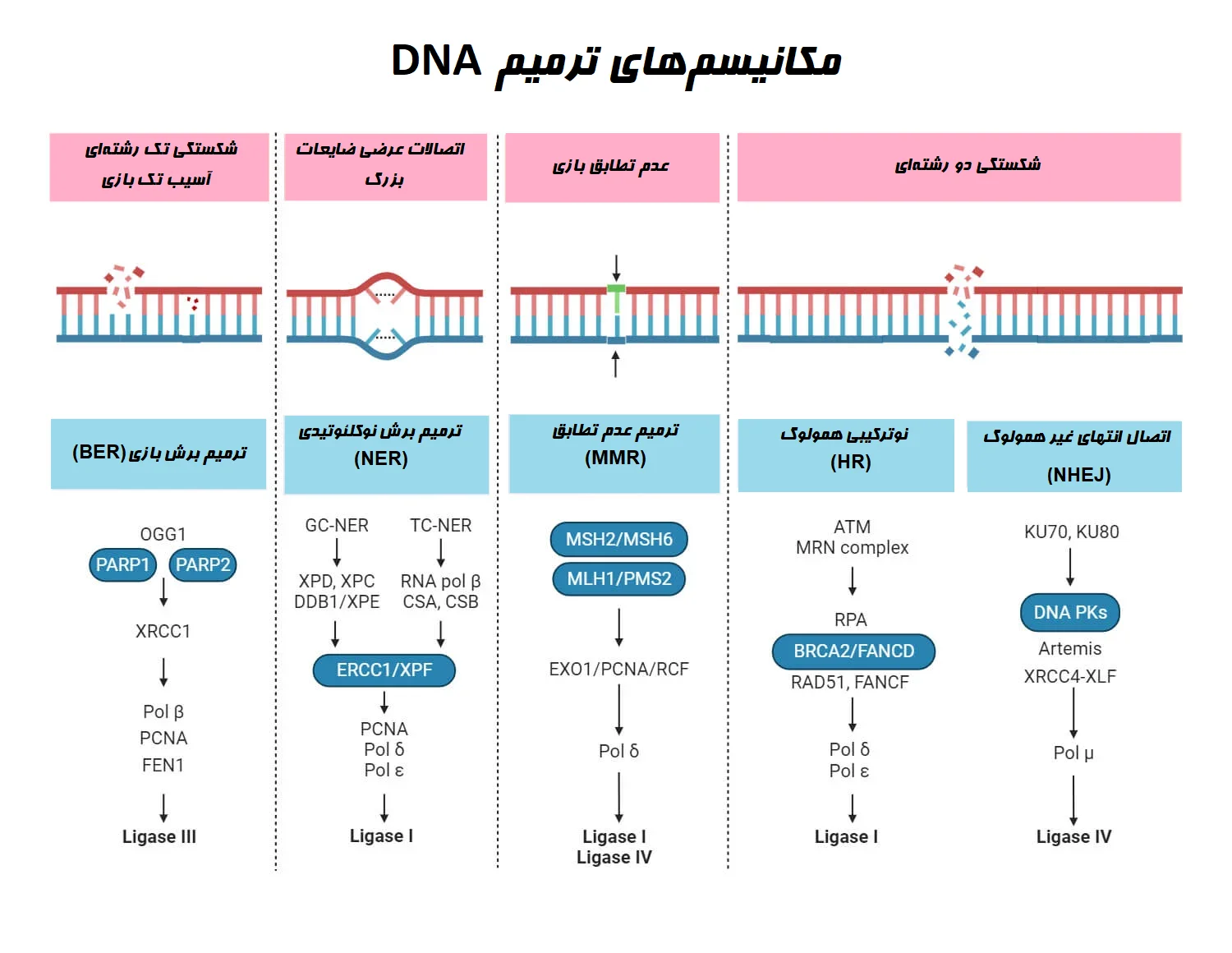
انواع و مکانیسمهای ترمیم DNA
مسیرهای مختلفی برای ترمیم DNA وجود دارد. این مسیرها شامل برگشت مستقیم (Direct reversal)، ترمیم برش (Excision repair)، بازسازی نابرابر (Mismatch repair) و ترمیم شکستگیهای DNA میباشد.
-
ترمیم برگشت مستقیم (Direct reversal repair)
- ترمیم برگشت مستقیم یک مکانیسم ترمیم DNA است که به طور مستقیم انواع خاصی از آسیب DNA را بدون نیاز به برش یا جایگزینی رفع میکند.
- دو نمونه از آسیب DNA که قابل برگشت هستند، ضایعات ناشی از اشعه ماوراء بنفش و بازهای آلکیله شده هستند.
- ضایعات ناشی از اشعه ماوراء بنفش که توسط نور UV ایجاد میشوند، میتوانند از طریق فرآیندی به نام فعالسازی مجدد نوری (Photoreactivation) که از انرژی نور مرئی برای شکستن ساختار DNA آسیب دیده استفاده کرده و بازهای پیریمیدین اصلی را بازیابی میکند، برطرف شوند.
- بازهای آلکیله شده را میتوان با آنزیمهایی مانند O6– آلکیلگوانین- DNA آلکیلترانسفراز (O6– alkylguanine- DNA alkyltransferase (AGT)) و دیاکسیژنازهای (Dioxygenase) مرتبط با AlkB که به ترتیب گروه آلکیل را حذف یا اصلاح میکنند، ترمیم کرد.
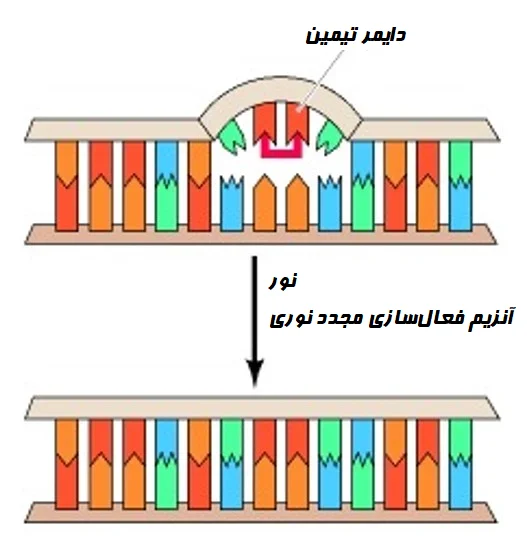
-
ترمیم برش بازی
- ترمیم برش بازی (Base excision repair (BER)) یک مکانیسم ترمیم DNA است که بازهای آسیب دیده را حذف و جایگزین میکند.
- این ترمیم شامل فعالیت DNA گلیکوزیلازهای مختلف مانند 8- اکسوگوانین DNA گلیکوزیلاز (8-oxoguanine DNA glycosylase (OGG1)) است. این آنزیمها بازهای آسیب دیده را شناسایی و حذف میکنند.
- BER هم شامل ترمیم پچ کوتاه (Short patch repair) میشود یعنی جایی که یک سایت آباسیک توسط آنزیمهای خاص پردازش و پر میشود، و هم ترمیم پچ بلند (Long patch repair)، که در آن شکافها متناسبسازی شده و سنتز DNA و به دنبال آن بستن (Ligation) انجام میشود.
- یکی از نمونههای BER، ترمیم DNA حاوی اوراسیل (Uracil) است. در این فرآیند یک DNA گلیکوزیلاز، باز اوراسیل را شناسایی و حذف و شکافی در DNA به نام سایت AP ایجاد میکند. سپس این شکاف توسط آنزیمی به نام AP اندونوکلئاز (AP endonuclease) بریده میشود. پس از آن، قند باقی مانده برداشته شده و شکاف با استفاده از DNA پلیمراز پر و با لیگاز (Ligase) مسدود میشود.
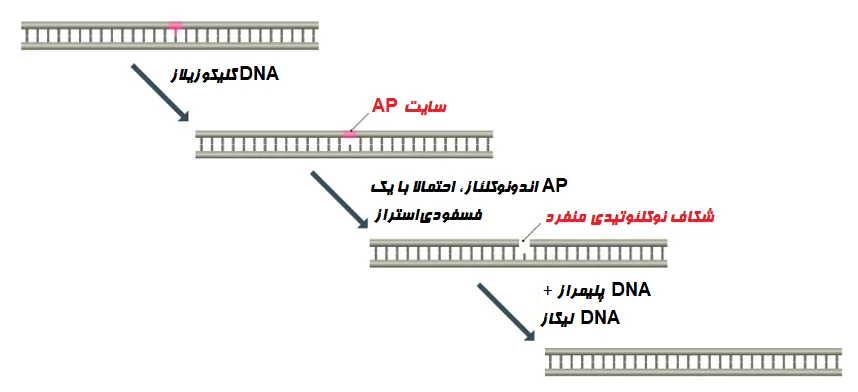
-
ترمیم برش نوکلئوتیدی

ترمیم برش نوکلئوتیدی
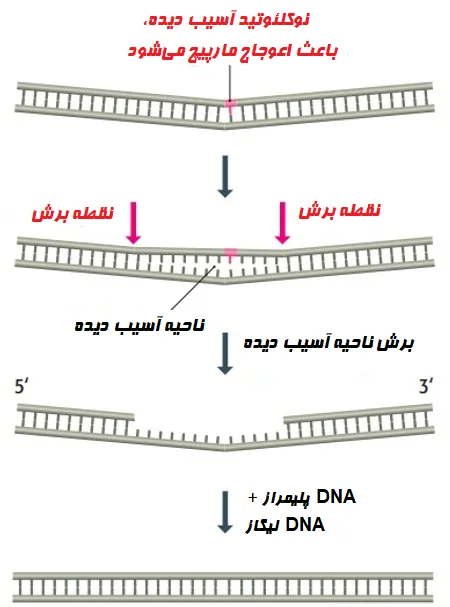
- ترمیم برش نوکلئوتیدی (Nucleotide excision repair (NER)) با ترکیبات افزایشی حجیم و ضایعات اتصال عرضی که از اشعه ماوراء بنفش یا قرار گرفتن در معرض مواد شیمیایی ناشی میشوند، سروکار دارد.
- NER قطعهای از نوکلئوتیدهای حاوی ضایعه آسیب دیده را حذف کرده و یک رشته DNA جدید را با استفاده از رشته آسیب ندیده به عنوان الگو، سنتز میکند.
NER از دو مسیر تشکیل شده است:
- ژنوم جهانی NER (Global Genome NER (GG-NER)) آسیبهای بزرگ را در کل ژنوم، از جمله مناطقی که به طور فعال رونویسی نشدهاند، ترمیم میکند.
- NER همراه با رونویسی (Transcription-Coupled NER (TC-NER)) آسیبهایی را که در رشته DNA رونویسی شده رخ میدهد، ترمیم میکند.
- جهش در ژنهای مسیر NER میتواند منجر به اختلالاتی مانند خشکپوستی رنگدانهای یا گزرودرما پیگمنتوزوم (Xeroderma pigmentosum (XP)) و برخی دیگر از بیماریهای تخریبکننده عصبی شود.
-
بازسازی نابرابر

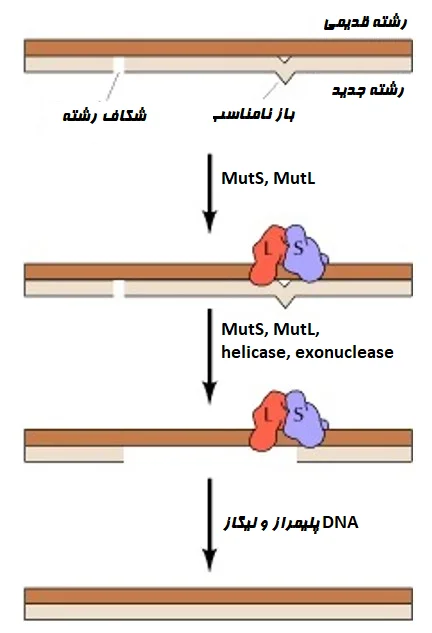
- مسیر بازسازی نابرابر (Mismatch repair (MMR))، عدم تطابق باز و لوپهای درج-حذف (Insertion-deletion loop) را که در حین فرایند همانندسازی رخ میدهد، ترمیم میکند. اکثر این خطاها با فعالیت تصحیح (Proofreading) DNA پلیمراز در طول فرایند همانندسازی برطرف میشوند، اما ممکن است برخی از آنها از قلم افتاده که باید بعداً اصلاح شوند.
- مسیر MMR شامل سه مرحله است: تشخیص عدم تطابق یا mismatch، تخریب رشته حاوی خطا، و سنتز توالی DNA صحیح.
- ابتدا کمپلکسهای پروتئینی مانند MSH2-MSH6 در پروتئین MutS، خطاهای عدم تطابق را پیدا میکنند و یک کمپلکس با MutL تشکیل داده که به ترمیم بیشتر کمک میکند. MutS و MutL کمپلکسهای پروتئینی مهم در یوکاریوتها (Eukaryote) هستند. در coli، پروتئین دیگری به نام MutH نیز نقش مهمی در ترمیم عدم تطابق دارد.
- در مرحله بعد، اگزونوکلئاز 1 (exonuclease 1 (Exo1)) رشته حاوی خطا را تخریب میکند در حالی که پروتئین همانندسازی A (Replication protein A (RPA)) با اتصال به DNA در معرض تخریب از تخریب بیشتر DNA جلوگیری میکند.
- سپس DNA پلیمراز δ توالی صحیح را سنتز میکند. در نهایت، DNA لیگاز هر شکاف باقی مانده در DNA ترمیم شده را مسدود میکند.
- جهش در ژنهای MMR میتواند منجر به سندرم لینچ (Lynch) (یک بیماری ارثی که با افزایش خطر ابتلا به سرطانهای روده بزرگ، تخمدان و سایر سرطانها مرتبط است) شود.
-
ترمیم شکستگی تک رشتهای (SSBR)
- شکستگیهای تک رشتهای (SSBs) در DNA میتواند به دلیل آسیب اکسیداتیو، سایتهای آباسیک یا بروز خطاهایی در فعالیت آنزیم توپوایزومراز DNA رخ دهد.
- این شکستگیها میتوانند همانندسازی DNA را مختل کنند، رونویسی را متوقف و فرآیندهای سلولی را فعال کنند که این امر میتواند منجر به مرگ سلولی شود.
- پروتئینهای PARP1 برای محافظت از تک رشتهای که در معرض شکستگی قرار گرفته است، آن را میپوشانند و به عنوان یک سپر عمل میکنند.
- SSBR را میتوان از طریق مسیرهای مختلفی که قبلاً در بالا توضیح داده شد، از جمله ترمیم برش بازی، ترمیم برش نوکلئوتیدی و بازسازی نابرابر انجام داد.

-
ترمیم شکستگی دو رشتهای
شکستگیهای دو رشتهای (DSBs) در DNA را میتوان از طریق دو مسیر ترمیم کرد: نوترکیبی همولوگ (Homologous recombination (HR)) و اتصال انتهای غیر همولوگ (Non-homologous end joining (NHEJ)).
نوترکیبی همولوگ (HR)
- HR یک مسیر ترمیم دقیق است که به یک توالی DNA منطبق به عنوان یک الگو نیاز دارد.
- HR در درجه اول از کروماتید خواهر (Sister chromatid)، یک کپی از DNA آسیب دیده، برای ترمیم استفاده میکند.
- HR در فازهای S، G2 و M مربوط به چرخه سلولی، زمانی که کروماتیدهای خواهر وجود دارند، بیشترین فعالیت را دارد.
- فرآیند HR شامل ایجاد DNA تک رشتهای (ssDNA) با تجزیه یک رشته از شکسته DNA و پوشاندن آن با پروتئینهایی مانند RPA است. Rad51 جایگزین RPA شده و ssDNA را با یک الگوی DNA همولوگ برای ترمیم، جفت میکند.
اتصال انتهای غیر همولوگ (NHEJ)
- NHEJ یک مکانیسم ساده و پرکاربرد است که مستقیماً انتهای شکسته DNA را بدون نیاز به الگوی DNA همولوگ میبندد.
- این مکانیسم میتواند در طول چرخه سلولی رخ دهد.
- پروتئینهایی مانند Ku70/Ku80، DNA-PKcs و LIG4/XRCC4 در NHEJ نقش دارند. Ku70/Ku80 از انتهاهای DNA محافظت و از نوترکیبی جلوگیری میکند، در حالی که DNA-PKcs و LIG4/XRCC4 به اتصال انتهای DNA کمک میکنند.
- مسیر NHEJ سریعتر است، اما در مقایسه با HR میتواند بیشتر مستعد خطا باشد.
همچنین بخوانید:
- DNA لدر چیست؟ انواع و موارد استفاده
- اشکال مختلف DNA: فرم A، فرم B، فرم Z
- دیانای ژیراز (DNA Gyrase): تعریف، ساختار، واکنشها و مکانیسمها
- انگشت نگاری DNA چیست؟ اصول، روشها، کاربردها
- آنالایزر DNA: اصول کار، مراحل کار و موارد استفاده
مترجم: صادق حسینیکیا