


اطلاعات عمومی، ویکی ژن
همانندسازی نیمه حفاظتی DNA در پروکاریوتها و یوکاریوتها
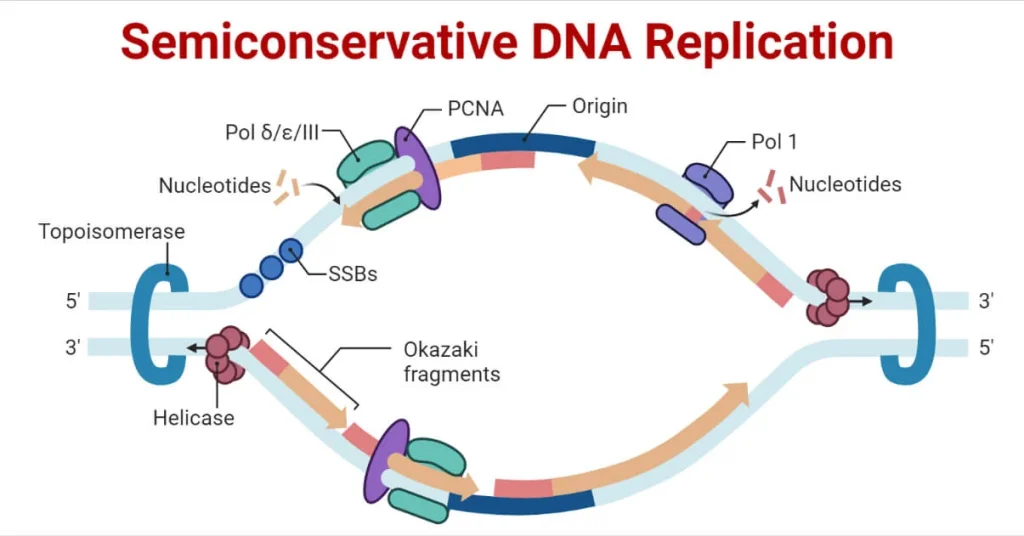
مقدمهای بر همانندسازی نیمه حفاظتی DNA
در مدل همانندسازی نیمه حفاظتی DNA، دو نسخه از مولکول DNA تولید میشود که هر نسخه حاوی یک رشته اصلی و یک رشته جدید سنتز شده است.
همانندسازی DNA نیمه حفاظت شده
- مدل DNA مارپیچ دوگانه واتسون و کریک، نوعی سیستم همانند سازی خودکار دارد.
- به دلیل اختصاصی بودن جفت بازها (آدنین به تیمین و گوانین به سیتوزین)، توالی بازها در زنجیره DNA، توالی زنجیره دیگر را تعیین میکند.
- این ویژگی باعث میشود که هر زنجیره به عنوان الگویی برای سنتز زنجیره دیگر عمل کند.
- واتسون و کریک برای همانندسازی DNA بیان کردند که همانندسازی شامل قطع پیوندهای هیدروژنی سپس پیچش و جدا شدن دو رشته پلی نوکلئوتیدی است.
- هر باز پورین یا پیریمیدین در هر رشته پلی نوکلئوتیدی، یک نوکلئوتید مکمل به سمت خود جذب میکند که برای پلیمریزاسیون در سلول به صورت آزاد موجود است و با پیوند هیدروژنی متصل میشوند.
نوکلئوتیدهای آزاد از طریق تشکیل پیوندهای فسفودی استری (اتصال بین دئوکسی ریبوزهای مجاور)، به محض اینکه در زنجیره الگوی اصلی قرار گرفتند، به یکدیگر متصل میشوند و زنجیره پلی نوکلئوتیدی جدیدی را تشکیل میدهند.
- نتیجه همانندسازی، تشکیل دو مولکول DNA دو رشتهای با توالی یکسان و مشابه با DNA اصلی است.
- یکی از رشتههای اصلی در یکی از این زنجیرههای دختری وجود دارد و رشته اصلی دیگر در زنجیره دختری دیگر وجود دارد.
- از آنجایی که هر مولکول DNAجدید، نیمی از مولکول DNA مادری را به همراه دارد و حفظ میکند، این الگوی همانندسازی نیمه حفاظت شده، نامیده میشود.
- همانند سازی DNA در پروکاریوتها و یوکاریوتها مشابه است اما یکسان نیست و در یوکاریوتها پیچیدهتر است.
علاوه بر حالت نیمه حفاظت شده، دو مدل دیگر در همانندسازی DNA وجود دارد که عبارتند از:
- همانندسازی حفاظت شده یا حفاظتی:
مولکول DNA جدید از دو رشته تازه سنتز شده تشکیل شده است و هر دو زنجیره در مارپیچ دوگانه مادری حفظ میشوند.
- همانندسازی پراکنده (غیر حفاظتی):
مارپیچ دوگانه مادری در طول همانندسازی شکسته میشود و قطعاتی از زنجیرههای اصلی با زنجیرههای تازه سنتز شده، مخلوط میشوند تا دو مارپیچ دوگانه جدید را تشکیل دهند.
آنزیمهایی که در همانندسازی DNA مشارکت دارند :
| آنزیم/پروتئین | عملکرد |
| DNA Helicase | مارپیچ DNA را با شکستن پیوند هیدروژنی بین بازهای نیتروژنی باز میکند |
| Topoisomerase | با ایجاد شکستگی مجدد در DNA به باز شدن پیچش در DNA کمک میکند |
| Single-strand binding (SSB) proteins | به DNA تک رشتهای متصل میشود تا از ایجاد پیچش جلوگیری کند. |
| Primase | آغازگرهای RNA مورد نیاز برای شروع همانندسازی را سنتز میکند |
| DNA pol I | فعالیت اگزونوکلئاز باعث حذف پرایمر RNA و DNA تازه سنتز شده جایگزین میشود |
| DNA pol II | نظارت بر جفت شدن صحیح بازها و تصحیح برعهده این آنزیم است |
| DNA pol III (α, δ and ε) | آنزیم اصلی که نوکلئوتیدها را به زنجیره اضافه میکند |
| Sliding Clamp | هنگام افزودن نوکلئوتیدها، به حفظ DNA پلیمراز کمک میکند |
| DNA Ligase | قطعات اوکازاکی را به هم متصل میکند تا زنجیره کامل ایجاد شود |
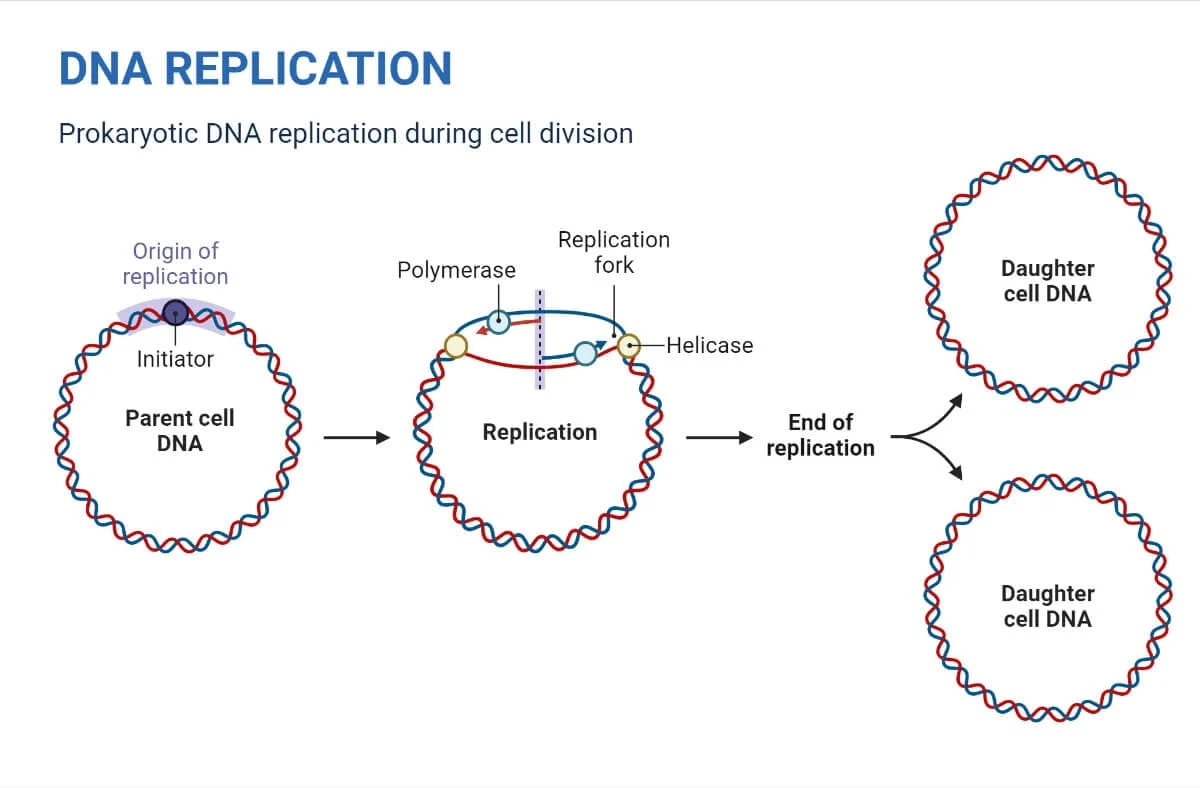
همانندسازی DNA در پروکاریوتها
مکانیسم تکثیر در پروکاریوتها را می توان در 3 مرحله طبقه بندی کرد:
- شروع
- طویل شدن
- خاتمه
شروع همانندسازی DNA در پروکاریوتها
- همانندسازی DNA از نقطه آغاز همانندسازی شروع میشود.
- ناحیه Ori تنها نقطه آغاز در همانندسازی است که در Ecoli یافت میشود (مانند اکثر پروکاریوتها).
- چندین توالی AT در این ناحیه وجود دارد که حدود 245 جفت باز طول دارد.
- ناحیه Ori دارای 13 و 9 جفت باز تکرار شونده است.
- کمپلکسی شامل 30 پروتئین DnaA ابتدا به 9 جفت باز تکرار شونده می چسبد و DNA را خم میکند تا مارپیچ DNA را در ناحیه تکرار شونده 13 جفت بازی، باز کند.
سپس پروتئینهای DnaC به DNA هلیکاز (DnaB) روی ناحیه شروع همانندسازی، کمک میکنند.
- DNA هلیکاز با قطع پیوندهای هیدروژنی بین جفت بازهای نیتروژنی، DNA را باز میکند. این فرآیند نیاز به هیدرولیز ATP دارد.
- ساختارهای Y شکل که به نام چنگالهای همانندسازی شناخته میشوند، با باز شدن DNA تشکیل میشوند.
- در ناحیه شروع همانند سازی، دو چنگال همانندسازی تشکیل میشود که با ادامه تکثیر به صورت دو جهته حرکت میکنند.
توپوایزومراز با ایجاد برش موقت در مارپیچ DNA و سپس مهر و موم مجدد آن، از پیچش مجدد در مارپیچ DNA هنگام باز شدن DNA ، قبل از چنگال همانندسازی، جلوگیری میکند.
- پروتئینهای اتصال تک رشتهای (SSB) تک رشتههای DNA را در نزدیکی چنگال همانندسازی میپوشانند تا از برگشت DNA تک رشتهای به داخل مارپیچ دوگانه جلوگیری کنند.
- DNA پلیمراز نمی تواند همانندسازی DNA را آغاز کند. فقط می تواند نوکلئوتیدها را در جهت 5′ به 3′ اضافه کند. به یک گروه 3′-OH آزاد نیاز دارد که بتواند با ایجاد یک پیوند فسفودی استر بین انتهای 3′-OH و 5′ فسفات نوکلئوتید بعدی، نوکلئوتیدها را اضافه کند. اگر یک گروه 3′-OH آزاد وجود نداشته باشد، نمی تواند نوکلئوتیدی اضافه کند.
- آنزیم RNA Primase یک قطعه RNA را که 5 تا 10 نوکلئوتید طول دارد و مکمل DNA الگو است، سنتز میکند و به عنوان پرایمر شناخته میشود زیرا این توالی سنتز DNA را آغاز میکند.
مرحله طویل شدن در همانندسازی DNA پروکاریوتی
- هنگامی که پرایمینگ کامل شد، DNA پلیمراز III در DNA شروع به کار میکند و افزایش طول شروع میشود.
- دو رشته DNA الگو خلاف جهت هم هستند، یک رشته در جهت ˈ5 به 3ˈ است و دیگری در جهت ˈ3 به ˈ5 است.
- تنها رشته DNA که میتواند به طور مداوم در جهت چنگال همانندسازی سنتز شود، رشتهای است که مکمل رشته DNA 3ˈ به ˈ5 است. این رشته که به طور مداوم سنتز میشود، تنها به یک پرایمر نیاز دارد.
- رشته دیگری که از چنگال تکثیر فاصله دارد، در قطعاتی به نام قطعات اوکازاکی سنتز میشود و در نتیجه به پرایمرهای زیادی نیاز دارد. جهت این رشته DNA ˈ5 به 3ˈ است.
- پروتئین حلقهای شکلی، DNA پلیمراز را در جای خود نگه میدارد و نوکلئوتیدها را اضافه میکند.
- در ادامه سنتز، پرایمرهای RNA با DNA جایگزین میشوند.
- با افزودن نوکلئوتیدهای DNA، فعالیت اگزونوکلئاز DNA Pol I باعث حذف پرایمرها میشود.
- شکاف بین دو قطعه DNA توسط DNA لیگاز ایجاد میشود که به تشکیل پیوندهای فسفودی استر کمک میکند.
- دو چنگال همانندسازی که در ناحیه شروع همانندسازی تشکیل شدهاند، در جهت عکس، به شکل دایره حرکت میکنند.
خاتمه همانند سازی DNA در پروکاریوتها
- مرحله خاتمه زمانی اتفاق میافتد که دو چنگال به هم میرسند و با هم ترکیب میشوند و دو مولکول DNA دو رشتهای مجزا تولید میکنند.
- پروتئین خاتمه همانندسازی (Tus)، با شناسایی توالی خاتمه (ter)، به DNA متصل میشود تا چنگالهای همانندسازی را که از یک جهت نزدیک میشوند و نیز حرکت DNA هلیکاز را مسدود کنند.
- چنگال دیگر نیز زمانی که با چنگال اول برخورد کند، متوقف میشود.
- کروموزومهای حلقوی باکتریایی میتوانند در حین تکثیر در هم بپیچند یا با پیوند کووالانسی به هم بپیوندند.
- توپوایزومراز IV حلقههای درهم تنیده را از بین میبرد.
- در نهایت، دو نسخه DNA در تقسیم سلولی به دو سلول جدید منتقل میشود.
بازخوانی و تصحیح DNA تکثیر شده
- شامل اسکن انتهای زنجیرههای DNA تازه تکثیر شده برای تصحیح خطاهای همانندسازی DNA است.
- این فرآیند با فعالیت اگزونوکلئاز 3ˈ به ˈ5 DNA پلیمراز انجام میشود.
- نوکلئوتیدهای نادرست را در انتهای ˈ3 حذف میکند تا نوکلئوتید صحیح با فعالیت پلیمراز ˈ5 به ˈ3 اضافه شود.
- در باکتریها، هر سه DNA پلیمراز (I، II و III) توانایی تصحیح را دارند.
- هنگامی که یک جفت باز، نادرست تشخیص داده میشوند، DNA پلیمراز جهت خود را با یک جفت باز DNA معکوس میکند و باز نادرست را جدا میکند. پس از برداشتن باز، پلیمراز می تواند باز صحیح را دوباره وارد کند و همانندسازی ادامه مییابد.
- میزان تصحیح در همانندسازی DNA میزان جهش را تعیین میکند.
شواهدی برای همانندسازی نیمه حفاظتی DNA در پروکاریوتها
Meselson و F.W. Stahl در سال 1958، با استفاده از DNA نشاندار شده (به صورت ایزوتوپی) و یک نوع تکنیک سانتریفیوژ (گرادیان چگالی) ، ماهیت نیمه حفاظت شده همانندسازی DNA را با موفقیت نشان دادند.
- چندین نسل از E.coli در محیطی حاوی NH4Cl با 15N کشت شد.
- هنگامی که DNA از این سلولها جدا شده و بر روی نمک کلرید سزیم (CsCl) سانتریفیوژ میشود، زمانی که چگالی آن برابر با محلول نمک میشود، DNA جدا میشود.
- DNA سلولهای کشت شده در محیط 15N تراکم بالاتری نسبت به DNA سلولهای کشت شده در محیط استاندارد 14N داشت.
- سلولهای E. coli که فقط 15N در DNA خود داشتند به محیط 14N منتقل شدند و تقسیم شدند.
- افزایش تقسیم سلولی با شمارش سلولی میکروسکوپی و با استفاده از سنجش کلنی مشاهده شد.
- DNA به صورت دورهای استخراج شد و با DNA خالص 14N و 15N مقایسه شد.
- چگالی DNA پس از یک تکرار متوسط بود.
- همانندسازی نیمه حفاظت شده نادیده گرفته شد زیرا DNA با چگالی بالاتر و کمتر تولید نمیکرد. نتیجه با هر دو تکرار نیمه حفاظت شده و پراکنده مطابقت داشت.
- همانندسازی نیمه حفاظتی DNA دو رشتهای با یک رشته DNAبا 15N و دیگری با 14N تولید میکند، در حالی که همانندسازی پراکنده DNA دو رشتهای را با هر دو رشته دارای مخلوطی از 15N و 14N تولید میکند. هر دوی اینها با چگالی متوسط ظاهر میشدند.
- DNA سلولها پس از دو تکرار، حاوی مقدار مساوی DNA با دو چگالی متفاوت بود که یکی از آنها با چگالی متوسط تنها برای یک تقسیم در محیط 14N و دیگری با DNA سلول هایی که منحصراً در محیط کشت 14N رشد کرده بودند، مطابقت داشت.
- این نتیجه با همانندسازی پراکنده متناقض بود و با فرضیه تکرار نیمه حفاظت شده مطابقت داشت.
همانندسازی DNA در یوکاریوتها
فرآیند تکثیر در یوکاریوتها را میتوان در 3 مرحله اصلی خلاصه کرد:
- شروع
- طویل شدن
- خاتمه
شروع تکثیر DNA در یوکاریوتها
- کروموزوم یوکاریوتی دارای چندین ناحیه شروع برای تکثیر است. انسانها میتوانند تا 100000 ناحیه شروع تکثیر داشته باشند.
- در مبدأ همانند سازی، یک کمپلکس پیش همانندسازی با سایر پروتئینهای آغازگر ساخته میشود. شامل:
- شش پروتئین برای تشخیص مبدا (ORC) (ORC1-6)
- پروتئین کنترل تقسیم سلولی (Cdc6).
- پروتئين تنظیمی کروماتین و عامل همانندسازی DNA (Cdt1).
- شش پروتئین کوچک نگهدارنده کروموزوم (MCM) (MCM2-7)
- این فرآیند در فاز G1 چرخه سلولی شروع میشود یعنی زمانی که موتورهای DNA هلیکاز همانندسازیکننده (MCM2-7 ) ، برای هر ناحیه آغاز به کار گرفته میشوند.
- کینازهایی که پس از ایجاد pre-RC شروع به فعال سازی کمپلکس میکنند. عبارتند از:
- کیناز وابسته به سیکلین (CdK)
- کیناز وابسته به Dbf4 (DdK)
- DdK و CdK پروتئین دیگری به نام Cdc45 را به کار میگیرند که تمام پروتئینهای تکثیر کننده DNA را فعال میکند.
- Cdc45، MCM2-7 و مجموعه چهار زیر واحدی Go-Ichi-Ni-San (GINS) برای تشکیل CMG هلیکاز فعال در طول فاز S ترکیب میشوند.
- CMG هلیکاز ناحیه شروع همانندسازی را از هم باز میکند.
- ساختارهای Y شکل که به نام چنگالهای همانندسازی شناخته میشوند، با باز شدن DNA تشکیل میشوند.
- در ناحیه شروع همانند سازی، دو چنگال همانندسازی تشکیل میشود که با ادامه تکثیر به صورت دو جهته پیشروی میکنند.
- DNA پیچ خورده زمانی تشکیل میشود که فعالیت DNA هلیکاز از نظر توپولوژیکی به رشته باز شده، فشار وارد کند که به دلیل ارتباط فاکتور همانندسازی A (RF-A) و توپوایزومراز با تغییر توپولوژی DNA در چنگال همانندسازی رخ میدهد.
- پروتئینها تک رشتههای DNA را در نزدیکی چنگال همانندسازی اشغال میکنند تا از برگشت DNA در حالت تک رشتهای به حالت مارپیچ دوگانه جلوگیری کنند.
- سنتز پرایمر توسط Primase انجام میشود که به شدت با DNA پلیمراز α مرتبط است.

مرحله طویل شدن در همانندسازی DNA یوکاریوتی
هنگامی که کمپلکس آغازگر تشکیل شد و سلولها به فاز S رفتند، کمپلکس تبدیل به replisome میشود.
- کمپلکس replisome یوکاریوتی، مسئول هماهنگی همانندسازی DNA است.
- DNA پلیمراز ε رشته الگو را به طور پیوسته سنتز میکند زیرا در همان جهتی است که DNA باز میشود.
- DNA پلیمراز α 20-30 نوکلئوتید را در طول قطعه Okazaki سنتز میکند، که توسط DNA پلیمراز δ روی رشته الگوی مخالف، به صورت ناپیوسته گسترش مییابد.
- DNA پلیمراز در جای خود نگه داشته میشود.
- RNase H پرایمر RNA را حذف میکند، اما یک ریبونوکلئوتید به انتهای 3′ قطعه Okazaki متصل باقی میماند.
- فلپ اندونوکلئاز 1 (FEN 1)، ریبونوکلئوتید باقیمانده را حذف میکند.
- پس از حذف پرایمر، شکاف بین قطعات اوکازاکی توسط DNA پلیمراز δ ازبین میرود.
خاتمه همانند سازی DNA در یوکاریوتها
- مرحله خاتمه زمانی رخ میدهد که دو چنگال همانندسازی به هم برسند. این محل معمولاً از پیش تعیین نشده است.
- دو رپلیزوم در جهت مخالف از یکدیگر عبور میکنند، و تصور میشود که کمپلکس CMG روی آخرین قطعه اوکازاکی می لغزد.
- یک مرحله مهم در خاتمه تکثیر یوکاریوتی، حذف CMG هلیکاز از کروماتین است.
- CMG هلیکاز با ubiquitination MCM7 حذف میشود و سپس Cdc48 وارد میشود و replisome را از بین میبرد.
- کروموزومهای یوکاریوتی خطی هستند، بنابراین همانندسازی DNA به انتهای کروموزوم نمیرسد.
- از آنجایی که قطعات اوکازاکی به پرایمرهای RNA نیاز دارند بنابراین منجر به از دست دادن DNA در هر چرخه میشود.
- سلولهای یوکاریوتی با استفاده از نواحی تلومر (توالیهای نوکلئوتیدی تکراری که هیچ ژنی در انتهای کروموزومها کد نمیکنند)، مشکل کوتاه شدن DNA در طول همانندسازی را حل میکنند.
- در انسان این توالی تکراری TTAAGGG است. این توالی 100 تا 1000 بار در نواحی تلومر تکرار میشود و از آسیب رسیدن به انتهای کروموزوم جلوگیری میکند.
- بنابراین تنها DNA ای که از دست میرود، قطعه تلومر است.
- در برخی از انواع سلولهای بنیادی و گلبولهای سفید، آنزیم تلومراز با گسترش توالی تلومر از تخریب سلولی جلوگیری میکند.
- این اتفاق در سلولهای سوماتیک رخ نمیدهد، بنابراین تلومرهای آنها با هر دور تکرار کوتاه میشود.
- هنگامی که تلومراز به اشتباه در سلولهای سوماتیک فعال میشود، میتواند منجر به سرطان شود.
تصحیح همانندسازی
- سه پلیمراز Pol α، Pol δ، و Pol ɛ ، ژنوم یوکاریوتی را تکثیر میکنند.
- Pol α به سنتز حدود 1.5 درصد از ژنوم یوکاریوتی کمک میکند و صدها عدم تطابق را در طول هر بار تصحیح، تشخیص میدهد.
- تصحیح اگزونوکلئولیتیک Pol δ خطاهای تشخیص داده شده توسط Pol α را تصحیح میکند.
- خطاهای وابسته به Pol δ و Pol α، توسط Pol ɛ رفع نمیشوند. با این حال، Pol δ میتواند خطاهای معرفی شده توسط Pol ɛ در رشته DNA الگو را تصحیح کند.
شواهدی برای همانندسازی نیمه حفاظتی DNA در یوکاریوتها
- J.H. Taylor و P. Woods در سال 1957 با شکافتن سلولهای نوک ریشه لوبیا (Vicia faba) و استفاده از تکنیکهای اتورادیوگرافی و میکروسکوپ نوری، شواهدی را دال بر همانند سازی نیمه حفاظت شده DNA در یوکاریوتها ارائه کردند.
- به سلولهای نوک ریشه Vicia faba تیمید رادیواکتیو تیمیدین (3H) برای علامت گذاری در مولکول DNA داده شد. آنها متعاقباً در محیطهای معمولی و بدون علامت با کلشی سین رشد کردند تا خصوصیات نیمه حفاظت شده در همانندسازی کروموزوم را مشاهده کنند.
- برای جلوگیری از جدا شدن کروماتید خواهری در طول آنافاز، کلشی سین اضافه شد.
- در نسل اول، رشته DNA اصلی با یک برچسب 3H در کنار یک رشته DNA بدون برچسب دیگر مشاهده شد، زیرا رادیواکتیویته به طور مساوی در هر دو رشته توزیع شده است.
- تنها یک رشته از بخش دوم DNA دارای رادیواکتیویته بود.
- این نتیجه، ماهیت نیمه حفاظت شده DNA را نشان داد.
تفاوت بین همانندسازی پروکاریوتی و یوکاریوتی
| ویژگی | پروکاریوت | یوکاریوت |
| ناحیه شروع همانندسازی |
تنها یک مبدا همانند سازی در هر مولکول DNA |
دارای چندین ناحیه شروع همانندسازی در هر کروموزوم |
| محل همانندسازی | سیتوپلاسم | هسته |
| نرخ همانندسازی | (سریع)1000 nucleotides/s | (آرام)50 to 100 nucleotides/s |
| DNA polymerase نوع | 5 | 14 |
| Telomerase | ندارد | دارد |
| حذف پرایمر RNA | DNA pol I | RNase H |
| طویل شدن رشته | DNA pol III | Pol α, pol δ, pol ε |
| Sliding clamp (گیره) | Sliding clamp | PCNA |
همچنین بخوانید:
- همانندسازی DNA-دوبله ژنیران
- همانندسازی ژن چیست؟
- DNA پلیمراز چیست؟
- آسیب DNA و ترمیم DNA: انواع و مکانیسم
مترجم: معصومه قریبی ششده